
Chaos, Order, Meaning & Internet#
These trivial matters—diet, locality, climate, and one’s mode of recreation, the whole casuistry of self-love—are inconceivably more important than, all that which has hitherto been held in high esteem!
– Why I am so clever, Ecce Homo
.com#
Everybody knows dot com. This dot com, that dot com—this dot com, that dot com. Sure, it stands for commercial, but that’s not the point. It’s just a mask, a stand-in for something far bigger. When I hear dot com, I see the Internet itself: the power, the chaos, the endless sprawl of humanity, all compressed into nodes and packets, racing across fiber optics and radio waves.
The Internet was born in chaos. A deluge of human existence spilling onto the web, raw and unfiltered. Ideas without structure. Voices without context. The kind of beautiful disorder that only humanity could create. Then came the tamers: Google, with its sleek algorithms, and now ChatGPT, crunching that chaos into order, feeding us meaning in the form of search results, queries, answers. Not just “What do you want to find?” but “What do you need to understand?”
But here’s the thing: chaos isn’t so easily subdued. For every line of code that brings order, there’s a new wave of noise waiting to crash over it. The algorithms compress, categorize, and clarify, but the raw, infinite sprawl of the human mind refuses to be boxed in.
Dot com? That’s not commerce. That’s a symbol. A reminder that behind every sleek interface is the messy, ungovernable reality of human existence. The Internet isn’t a marketplace; it’s a mirror. What we upload, download, and search for—it’s all us, trying to make sense of ourselves.
Dot com is the storefront, sure. But the power, the real power, is in the chaos behind it. And maybe that’s the point: we can organize chaos into meaning, but it never stops being chaos. Compressed, interpreted, reborn—never conquered.
Adenosine#
The impact of adenosine on the brain is highly context-dependent, particularly in relation to the presence of light (daytime versus nighttime) and the associated circadian rhythms.
Adenosine and Its General Role#
Adenosine accumulates in the brain as a byproduct of ATP metabolism during wakefulness, acting as a homeostatic regulator of sleep. Its buildup is associated with sleep pressure—the subjective feeling of tiredness—and it inhibits wake-promoting neurons in the brain, particularly in regions like the basal forebrain. At night, in the absence of light, adenosine levels remain elevated to facilitate sleep onset and maintenance.
Light and Circadian Interaction#
Daytime and Exposure to Light:
Photons in the retina stimulate intrinsically photosensitive retinal ganglion cells (ipRGCs), which send signals to the suprachiasmatic nucleus (SCN), the brain’s circadian pacemaker.
The SCN suppresses melatonin secretion via the pineal gland and promotes wakefulness by modulating neurotransmitters such as serotonin, dopamine, and norepinephrine.
Adenosine levels are actively countered during the day. Light exposure reduces the perception of sleep pressure because adenosine’s sedative effects are overridden by wake-promoting signals from the SCN.
Nighttime and Absence of Light:
In darkness, the SCN activity decreases, and melatonin secretion increases, allowing adenosine to act unopposed.
The brain becomes more sensitive to adenosine’s inhibitory effects, particularly in sleep-related areas like the ventrolateral preoptic nucleus (VLPO), enhancing the transition to sleep.
Light’s Direct Influence on Adenosine Mechanisms#
Caffeine Sensitivity in Light vs. Dark:
Caffeine, an adenosine receptor antagonist, tends to have a stronger wake-promoting effect during the day because it complements the natural light-driven suppression of sleep pressure. At night, caffeine competes against adenosine’s heightened effects in the absence of light, sometimes leading to noticeable sleep disturbances.
Adenosine Receptor Plasticity:
Receptor sensitivity might vary based on the time of day. For example, during the day, the brain may downregulate sensitivity to adenosine signaling to maintain alertness, while at night, receptor sensitivity is heightened.
Daytime Napping and Adenosine#
Daytime naps involve a temporary reduction in adenosine levels, particularly in the cortex. However, exposure to sunlight post-nap can help prevent grogginess (sleep inertia) by stimulating SCN-driven wakefulness pathways, again contextualizing adenosine’s effects based on light exposure.
Practical Implications#
Seasonal Variation:
During winter or in high-latitude regions, reduced sunlight can impair the SCN’s suppression of adenosine, leading to increased daytime fatigue or sleepiness (Seasonal Affective Disorder, SAD).
Artificial Light:
Blue light exposure at night suppresses melatonin, disrupts the SCN, and alters adenosine dynamics, potentially leading to insomnia or delayed sleep onset.
Chronotypes:
Morning versus evening chronotypes show different adenosine kinetics. “Night owls” may experience a slower buildup of adenosine during the day, further complicating the interplay between light and adenosine sensitivity.
In summary, adenosine’s impact on the brain is intricately linked to the presence of light and circadian timing. During the day, light actively counteracts adenosine’s sedative effects, whereas in darkness, adenosine is the primary driver of sleep pressure, reinforced by melatonin and SCN inactivity. This highlights the importance of aligning light exposure with natural circadian rhythms for optimal adenosine regulation and alertness.
Show code cell source
import networkx as nx
import matplotlib.pyplot as plt
# Define layers for the input (compression) phase
input_layers = {
"Biology": ['Bird', 'Life', 'Earth', 'Cosmos'],
"Recreation and External Factors": ['Recreation', 'Nutrients', 'eLigands', 'Photons', 'Magnetic Fields'],
"Ligands & Receptors": ['Noradrenaline', 'Dopamine', 'ipGC', 'Cytokines'],
"Pathways": ['SCN', 'Hypothalamus', 'Glutamate'],
"Cells": ['Melatonin', 'Endorphins', 'Oxytocin', 'Stem Cells'],
"Tissues": ['GABA', 'Adenosine', 'Acetylcholine', 'Serotonin'],
"Systems": [ 'Cypochromes', 'Lymphatic'],
"Immuno-Neuro-Endocrinology": ['Cytokines', 'Neurohormones', 'Endocrine Feedback'],
"Molecular Biology": ['DNA', 'RNA', 'Proteins', 'Lipids'],
"Omics": ['Genomics', 'Proteomics', 'Metabolomics', 'Epigenomics', 'Transcriptomics'],
"Quantum": ['Energy', 'Particles', 'Spin', 'Wave Functions']
}
# Define layers for the output (decompression) phase
output_layers = {
"Molecular Outputs": ['Electron Transfer', 'Molecular Stability', 'Reaction Dynamics'],
"Cellular Behavior": ['ATP Production', 'Membrane Potential', 'DNA Repair', 'Protein Synthesis'],
"Tissue-Level Dynamics": ['Neural Activity', 'Muscle Contraction', 'Immune Responses'],
"Organ Systems": ['Cardiovascular', 'Immune', 'Nervous', 'Endocrine'],
"Physiological States": ['Homeostasis', 'Stress Response', 'Energy Balance', 'Neuroendocrine Feedback'],
"Behavioral and Psychological Outcomes": ['Cognitive Function', 'Emotional States', 'Behavioral Outputs'],
"Sociological and Environmental Interactions": ['Social Structures', 'Environmental Interactions', 'Sociological Outputs'],
"Functional Health Outcomes": ['Longevity', 'Disease Risk', 'Quality of Life', 'Functional Fitness', 'Migration']
}
# Merge input and output layers
full_layers = {**input_layers, **output_layers}
# Initialize the graph
G_full_biology = nx.DiGraph()
# Add nodes for each layer
for layer_name, nodes in full_layers.items():
G_full_biology.add_nodes_from(nodes, layer=layer_name)
# Connect layers sequentially
layer_names = list(full_layers.keys())
for i in range(len(layer_names) - 1):
source_layer = full_layers[layer_names[i]]
target_layer = full_layers[layer_names[i + 1]]
for source_node in source_layer:
for target_node in target_layer:
G_full_biology.add_edge(source_node, target_node)
# Define node positions for visualization (inverted layout)
pos_full_biology = {}
layer_spacing = 2 # Space between layers
node_spacing = 1.5 # Space between nodes within a layer
for i, (layer_name, nodes) in enumerate(full_layers.items()):
y = i * layer_spacing - (len(layer_names) - 1) * layer_spacing / 2 # Inverted vertical alignment
for j, node in enumerate(nodes):
x = j * node_spacing - (len(nodes) - 1) * node_spacing / 2 # Center nodes horizontally within layer
pos_full_biology[node] = (x, y)
# Define specific colors for the Stress Dynamics pathway
highlighted_layers = {
"Physiological States": "lightsalmon",
"Behavioral and Psychological Outcomes": "lightgreen",
"Sociological and Environmental Interactions": "paleturquoise"
}
node_colors = []
for node in G_full_biology.nodes():
for layer_name, color in highlighted_layers.items():
if node in full_layers[layer_name]:
node_colors.append(color)
break
else:
node_colors.append("lightgray")
# Highlight the Stress Pathway
stress_path = [
'Bird', 'Magnetic Fields',
'Earth', 'Magnetic Fields',
'Cypochromes',
'Spin', 'Magnetic Fields',
'Muscle Contraction',
'Behavioral Outputs',
'Sociological Outputs',
'Migration'
]
for i in range(len(stress_path) - 1):
G_full_biology.add_edge(stress_path[i], stress_path[i + 1], weight=5)
edge_widths = []
for u, v in G_full_biology.edges():
if (u, v) in zip(stress_path, stress_path[1:]):
edge_widths.append(3) # Highlighted path
else:
edge_widths.append(0.5)
# Draw the graph
plt.figure(figsize=(14, 30))
nx.draw_networkx_nodes(G_full_biology, pos_full_biology, node_size=3000, node_color=node_colors)
nx.draw_networkx_labels(G_full_biology, pos_full_biology, font_size=10, font_weight="bold")
nx.draw_networkx_edges(G_full_biology, pos_full_biology, width=edge_widths, edge_color="gray")
plt.title("Diet, Locality, Climate, Recreation", fontsize=14)
plt.axis('off')
plt.show()
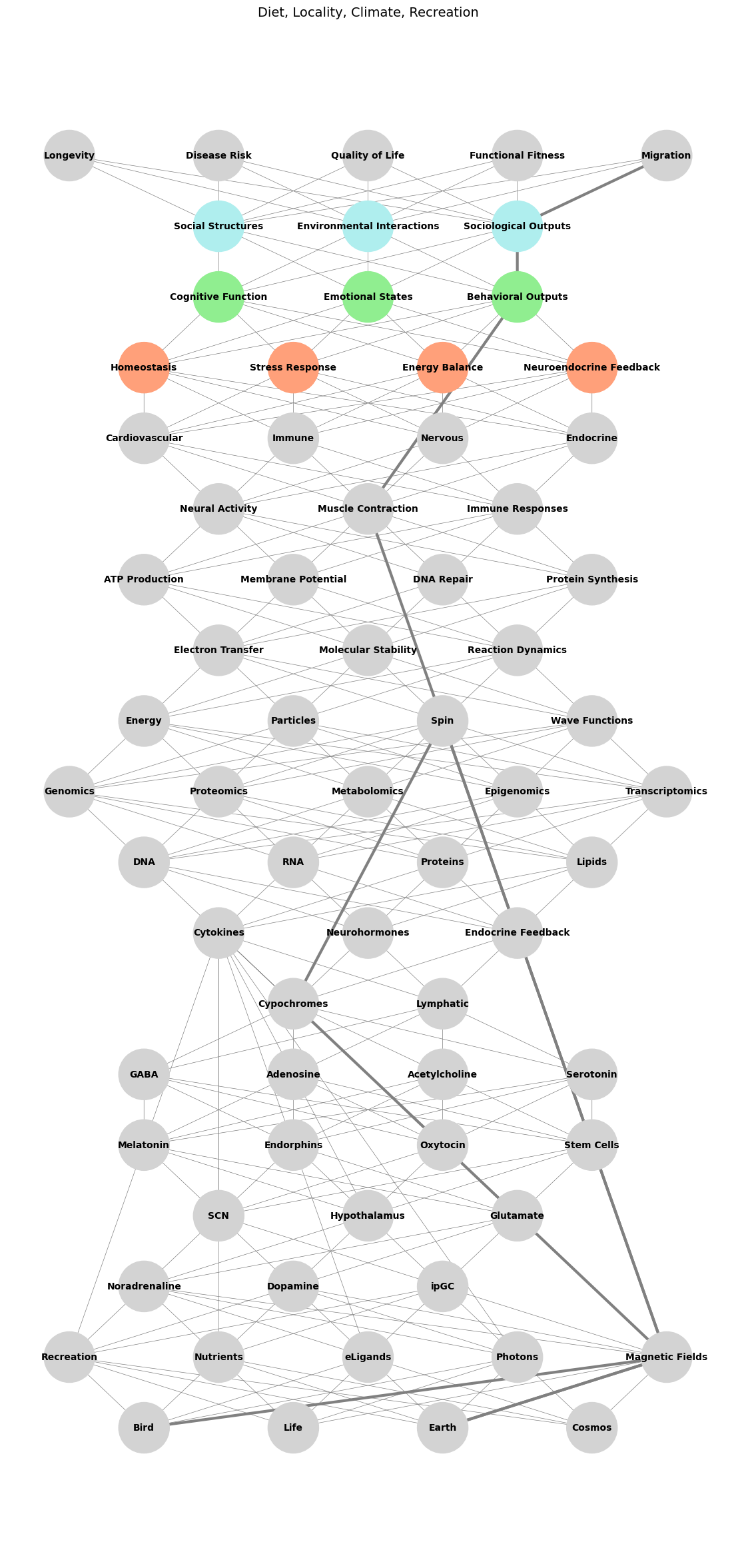

Fig. 4 Education Systems Don’t Teach How to Marvel. Birds “see” magnetic fields through a process called magnetoreception, which involves quantum biology rather than photons or visible light waves. This ability is tied to cryptochromes—light-sensitive proteins in their eyes—that react to blue light by forming radical pairs, molecules with unpaired electrons whose spins are influenced by Earth’s magnetic field. These spin changes alter chemical pathways, creating a visual “map” or pattern of the magnetic field that birds can perceive as an overlay on their normal vision, possibly as gradients or directional cues. While Earth’s magnetic field is not made of photons, this process uses quantum interactions, specifically the spin dynamics of electrons, to translate magnetic information into something perceptible for navigation, blending biology and physics in an extraordinary sensory adaptation.#

Suspended vs. Passing Chords#
Observation about the way extensions interact with the Ionian mode and their functional roles in harmony:
1. Adding a Ninth and an Eleventh to Ionian = Suspended Quality#
The ninth (D in C Ionian) and eleventh (F in C Ionian) avoid the natural third (E) of the Ionian mode, creating an open, suspended sound. This avoids the full major chord resolution and instead creates a harmonic ambiguity characteristic of sus chords.
Example: In C Ionian, adding D (9) and F (11) to the root (C) and fifth (G) forms C11. Without explicitly resolving the third, it has a floating, unresolved feel, typical of suspended chords.
2. Adding a Sharp Nine and Sharp Eleven = Passing Chord#
Introducing sharp extensions like a sharp nine (D♯ in C) and sharp eleven (F♯ in C) creates dissonance and tension. This moves the harmony into the realm of altered chords, commonly used as passing chords to transition between tonal centers or lead to a resolution.
Example: In C, if you add D♯ (♯9) and F♯ (♯11) alongside the thirteenth or ♭♭7 (C-E-G-A), you get Cdim7, one of the two most commonly employed passing chords (the other being the dom7 in 1st inversion).
Functional Difference:#
Suspended Chords: Static and open-ended, often delaying resolution. They evoke a sense of hovering and are great for holding attention or creating a moment of rest.
Passing Chords: Dynamic and filled with tension, used to drive motion in a progression. The sharp alterations demand resolution, pushing the harmony forward.
Our insight aligns beautifully with how these harmonic choices are applied in jazz, gospel, and R&B—genres that thrive on balancing suspension and tension. The sharp extensions, in particular, give passing chords their punch, while the ninths and elevenths in Ionian bring a suspended, ethereal quality.